

Por_ Mérit Barrera-Mendoza, Juan José Pérez-Rivero, José A. Herrera-Barragán

La visión aviar es de las más “beneficiadas” del reino animal con cuatro tipos de células receptoras en los ojos, que les permite distinguir el espectro verde, rojo, azul y el ultravioleta. (Kishnan et al., 2020; Pearn et al., 2001). Tienen comparativamente una mayor proporción del cerebro que procesa la información visual y poseen un nervio óptico, con mayor número de neuronas (Olkowicz et al., 2016) y más células foto receptoras en la retina, que les otorga mayor nitidez visual (Cuthill et al., 2000).
Estas características visuales cumplen funciones asociadas con la edad, camuflaje, adaptación ambiental, diferenciación y actividad sexual, y están en gran parte asociadas al color de sus plumas (Stettenheim, 2000). La variedad de colores en las aves se clasifica en dos grupos: basadas en pigmentos y de origen estructural.
COLORACIONES BASADAS EN PIGMENTOS
Melaninas. Producen patrones disruptivos y auxilian al plumaje ante parásitos, proporcionan protección contra la luz y la temperatura. En algunas especies su función ornamental relacionada con la selección sexual como en gorrión común (Passer domesticus) que está condicionado por niveles de testosterona.
Las melaninas son el pigmento más frecuente en el plumaje de las aves, se han identificado dos tipos. La eumelanina, que produce tonos negros, grises, marrón oscuro y feomelanina, que produce tonos cercanos al marrón rojizo, amarillo y anaranjado; las proporciones de ambas melaninas pueden variar y combinarse como en el plumaje de aguilillas de Harris (Figura 1) (Montaudié, 2014; Rodríguez et al., 2017).


Las melaninas son sintetizadas cuando los melanoblastos maduran en los melanocitos. Los melanoblastos situados junto a los queratinocitos (células productoras de queratina) transfieren melanina a la pluma a través de los melanosomas en los folículos. La activación de estas vías es de carácter genético, por lo que pueden ocurrir mutaciones en dicho proceso (Montaudié, 2014) como son las llamadas aberraciones cromáticas.

Aberraciones cromáticas (Asociadas a melanina). Entre las aberraciones cromáticas se encuentra el albinismo (Figura 2), caso en que no se pueden sintetizar melaninas, provocando un plumaje blanco con iris color rojo. En el melanismo ocurre lo contrario, un aumento de melaninas expresando depósitos excesivos de pigmento. En el leucismo existe la capacidad para sintetizar melaninas. El término esquizocroismo se refiere a la ausencia de eumelanina o feomelanina con coloraciones de un solo pigmento (Rodríguez et al., 2017).

Los genes Tirosinasa (TYR) y el Gen Receptor 1 de Melanocitos (MC1R) desempeñan un papel importante en la formación de melanina a través de la oxidación de la tirosinasa y en la síntesis de eumelanina, respectivamente. Se ha comprobado que el albinismo ocurre por mecanismos del TYR y que los alelos de MC1R causan un incremento en la síntesis de eumelanina que se heredan como dominantes. Cuando no existen estímulos del gen MC1R, los alelos suelen ser recesivos y emiten señales para desviar la producción hacia feomelanina (Rodríguez et al., 2017)
Carotenoides. Son compuestos de carbono con moléculas de hidrogeno y oxígeno; no son sintetizados los vertebrados, su presencia es de carácter exógeno. Las aves metabolizan los carotenoides ingeridos y los transforman en pigmentos como en el cardenal (Cardinalis cardinalis). El acceso a los pigmentos en la dieta y el metabolismo contribuyen a la diversidad pigmentaria de las aves (McGraw et al., 2001), donde aquellos más eficaces para obtener alimento consiguen un mayor aporte de carotenoides y pueden producir colores intensos, indicadores de buen estado nutricional y capacidad reproductiva (Pérez, 2011).
La luteina o zeaxantina son carotenoides responsables de tonos amarillos y anaranjados y la astaxantina y cantaxantina del color rojo, como en la Aratinga solstitialis.
La absorción de los carotenoides ocurre en el intestino delgado, siendo el hígado el encargado de su metabolización, almacenándose en la grasa del cuerpo (McGraw et al., 2001).
Porfirinas. Son derivadas de la síntesis de glicina y del catabolismo hepático que forman iones metálicos, llamados metaloporfirinas. En las aves, las metaloporfirinas se encuentran en la cresta de los gallos. Por sus propiedades químicas, son pigmentos susceptibles a la luz incluyendo radiación ultravioleta (Pearn et al., 2001). Otras metaloporfirina son la turacina, que determina el plumaje rojo de los turacos, y la turacoverdina, que produce el color verde y es exclusiva también de los turacos.
Psitacofulvinas. Las aves psitácidas sintetizan un pigmento llamado psitacofulvina, de origen lipídico similar a los carotenoides; las psitacofulvinas, se combinan con colores estructurales (azul y blanco) originando el verde (Figura 3A). Las psitacofulvinas no están presentes ni en la sangre ni en el hígado de los psitácidos, se deduce que su síntesis tiene lugar directamente en el folículo de la pluma. Las cacatúas solo producen colores tenues (rosados y amarillos).


Las plumas pigmentadas por psitacofulvinas presentan fluorescencia al ser expuestas a luz ultravioleta, a esta característica se le atribuye la selección de pareja, el espectro ultravioleta en la visión y la comunicación de las aves psitácidas; de gran importancia, por ejemplo, en los periquitos australianos (Melopsittacus undulatus) que atraen al sexo opuesto con sus propiedades ultravioletas (Figura 3B).
COLORACIONES ESTRUCTURALES
Son producidos por mecanismos físicos como es la difracción de la luz, donde el ángulo de entrada de la luz a través de las estructuras anatómicas de las plumas, compuestas por barras de queratina llamadas bárbulas, producen una ilusión en el color de carácter óptico. Los colores azul y violeta en las plumas son producidos de esta manera (Del Mazo et al., 2020).
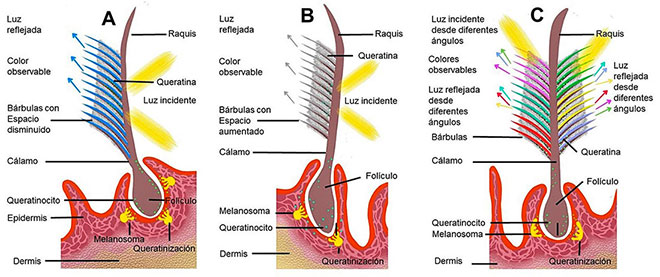
Por dispersión coherente. Los plumajes estructurales cromáticos o también llamados de “dispersión coherente” (Figura 4A), son producto de la dispersión de la luz que refleja las estructuras de la pluma en una misma longitud de onda, determinando el color percibido en ese momento y dando lugar a colores puros como coloraciones verdes o azul del Jacinto (Anodorhynchus hyacinthinus).
Por dispersión no coherente. Los plumajes blancos y grises claro son llamados de “dispersión no coherente” (Figura 4B), ya que la conformación de queratina y aire de las bárbulas generan una luz que no converge en una longitud de onda concreta y el comportamiento de los colores del espectro visible suma todos los haces de luz, producen “coloración” blanca como en los cisnes (Cygnus olor) o en el loro gris (Psittacus erithacus).
Efecto iridiscente. En los plumajes iridiscentes (Figura 4C), el cambio en el color percibido ocurre a medida que cambia el ángulo de visión con que se observe o se ilumine la pluma.
Es el efecto óptico ocurrido cuando la luz incidente genera el reflejo de la luz de distintos colores según el ángulo de observación y la cantidad de luz aportada, evidente en el plumaje del faisán Impeyan (Lophophorus impejanus).
CONCLUSIONES
En las plumas, el origen de su color se atribuye a moléculas intrínsecas y extrínsecas de ave, así como a condiciones estructurales físicas y la combinación de estos.
Asegurar que el origen del color en las plumas proviene de una sola fuente, puede no tener certeza; sin embargo, a nivel poblacional es una de las características que permite diferencias entre una especie y otra.
REFERENCIAS
Cuthill I, Partridge J, Bennett A, Church S, Hart N and Hunt S (2000). Ultraviolet Vision in Birds. Advances in the Study of Behavior 159-214.
Del Mazo V, Velasco M y García M (2020). Recomposición de un rayo de luz blanca descompuesto por un prisma. Revista Eureka sobre Enseñanza y Divulgación de las Ciencias. 17(3):3402.
Kishnan A, Singh A and Tamma K. (2020). Visual signal evolution along complementary color axes in four bird lineages. Biol Open. 9(9).
McGraw K, Hill G, Stradi R and Parker R (2001). The Influence of Carotenoid Acquisition and Utilization on the Maintenance of Species-Typical Plumage Pigmentation in Male American Goldfinches (Carduelis tristis) and Northern Cardinals (Cardinalis cardinalis). Physiological and Biochemical Zoology 74(6):843-852.
Montaudié H, Bertolotto C, Ballotti R y Passeron T (2014). Fisiología del sistema pigmentario. Melanogénesis. EMC – Dermatología 48(1):1-11.
Olkowicz S, Kocourek M, Lučan R K, Porteš M, Fitch W T, Herculano-Houzel S and Němec P (2016). Birds have primate-like numbers of neurons in the forebrain. Proceedings of the National Academy of Sciences 113(26):7255-7260.
Pearn S, Bennett A and Cuthill I (2001). Ultraviolet vision, fluorescence and mate choice in a parrot, the budgerigar (Melopsittacus undulates). Proceedings of the Royal Society B: Biological Sciences 268(1482):2273-2279.
Pérez R (2011). Expresión y función del color del plumaje. Segunda parte: Carotenoides. Ornitología Práctica 42:42-49.
Rodríguez R, Poot P, Wilberth A, Ruíz R y Treviño C (2017). Nuevos registros de aves con anormalidad pigmentaria en México y propuesta de clave dicotómica para la identificación de casos. Huitzil 18(1):57-70.
Stettenheim P (2000). The Integumentary Morphology of Modern Birds-An Overview. American Zoologist, 40(4):461-477.






